
Abstract
.
Objective: The in vivo nephroprotective effect of quercetin against sodium fluoride (NaF)-induced damage was studied. Methods: Renal injury was induced by daily administration of NaF (600 ppm) through drinking water for 1 week. The levels of reduced glutathione (GSH), lipid peroxidation as well as superoxide dismutase and catalase activity of kidney homogenates were determined. The serum markers of glomerular damage, including creatinine, serum urea, and blood urea nitrogen levels, were also assessed. Results: The study revealed that administration of fluoride resulted in a significant down-regulation of antioxidant defense coupled with an increased serum level of glomerular damage markers.
The administration of quercetin prior to fluoride reversed the antioxidant–oxidant balance to control (fluoride-untreated) level. The level of protection obtained for the 20 mg/kg quercetin treatment was equivalent to the positive control, ascorbic acid (10 mg/kg). The therapeutic implication of antioxidants in fluoride-induced nephrotoxicity is discussed.
.
Conclusions: This study showed that NaF intoxication caused renal damage by increasing oxidative stress, and quercetin and vitamin C administration gave protection against fluoride-induced oxidative stress to some degree.
..
Keywords: kidney, oxidative stress, quercetin, sodium fluoride, nephroprotection
.
INTRODUCTION
The natural source of fluoride for humans is from drinking water and food. However, recent studies estimated that about 20% of pharmaceuticals and 30–40% of agrochemicals are organofluorines.1Fluoride-containing compounds are also commonly used in household products and office (e.g., dental products, refrigerants, and fire extinguishers).2 Prolonged intake of fluoride or excessive consumption results in fluorosis, leading to a progressive degenerative disease, often characterized by dental mottling and skeletal dysfunctions including crippling deformities, osteoporosis, and osteosclerosis.1 Several studies revealed that fluoride intoxication in the hippocampus of the rat brain can lead to the degeneration of neuronal systems and an alteration of the oxidant–antioxidant balance in other organs and tissues.3 The kidney has a prominent role in fluoride metabolism as 50–80% of fluoride is removed via urinal excretion.4
Not surprisingly, the kidney is one of the major organs affected by fluoride intoxication, and numerous studies have established a close correlation between fluoride intake and renal injury. Hence, rats [See below], chronically intoxicated with sodium fluoride (NaF) have been shown to display histological renal changes, interstitial edema, tubular destruction, and glomerular and medullary hyperemia. Hand in hand with the typical kidney pathology, fluoride-intoxicated rats showed an increased rate of reactive oxygen species (ROS) generation and lipid peroxidation.5 Since antioxidants offer therapeutic values in reversing and/or preventing fluoride-mediated cell damage, several laboratories worldwide, including ours, are currently engaged in the search of potential remedies from synthetic and natural sources.
Polyphenolic compounds are the largest groups of secondary metabolites occurring throughout the plant kingdom. So far, over 8000 polyphenols, of which the largest group were the flavonoids, are known to be characterized.6 Structurally, flavonoids are made of two aromatic rings (ring A and ring B) linked through a three-carbon chain that usually forms an oxygenated heterocyclic ring (ring C). Variation in minor structural features (mainly in ring C) divided flavonoids into several families: flavonols, flavones, flavonols, isoflavones, anthocyanidin, and others. Many flavonoids have been shown to display antioxidant effects mainly through free radical scavenging and metal chelation.7 Quercetin (3,5,7,3′,4′-pentahydroxyflavone), one of by far the most known antioxidant flavonols, is known to occur in fruits and vegetables.8 In addition to radical scavenging and metal chelating effects, quercetin processes lipid peroxidation inhibitory effect.1
Some of the biological properties reported for quercetin include scavenging of superoxide anion in ischemia-reperfusion injury, protection against oxidative stress induced by ultraviolet light, for uses in spontaneous and experimentally induced hypertension, inhibition of secondary biliary cirrhosis, vasodilator effect, inhibition of low-density lipoprotein (LDL) oxidation, prevention of platelet aggregation and antiangiogenic effect, inhibition of carcinogenesis, inhibition of enzymes including tyrosine kinase and nitric oxide synthase, antiallergic and antiulcer effects, and anti-inflammatory property through NF-κB inhibition.8,9 The nephroprotective effect of quercetin on cisplatin-induced nephrotoxicity has been reported in recent years. Accordingly, Behling et al.9 revealed that the beneficial effect of quercetin was associated with its vasodilator properties, leading to an increase in renal blood flow. Similar studies by Sánchez-González et al.10 revealed that the beneficial effect of quercetin in cisplatin-induced nephrotoxicity is associated with inhibition of NF-κB-p65 activation, leading to a reduction of the inflammatory process. Various other studies have also demonstrated the beneficial effect of quercetin in reducing cisplatin-induced acute tubular necrosis and nephrotoxicity without compromising its antitumor effects.10,11 To date, the in vivo effect of quercetin on fluoride nephrotoxicity and redox parameters has not yet been established, and this study was primarily designed to address this key pharmacological parameters. It is also well known that vitamin C has a nephroprotective role in NaF-induced nephrotoxicity and oxidative stress.1 Therefore, vitamin C was used as a standard nephroprotective agent in this study.
.
MATERIALS AND METHODS
Animals
Male Wistar rats (Rattus norvegicus albinus) of 200–250 g body weight were used. The animals were housed in well-ventilated rooms, maintained at a temperature of 24 ± 2°C with a 12-h light/dark cycle, and 60 ± 5% humidity. Animals were fed with standard diet (supplied by Pasture Institute, Tehran, Iran) and water was provided ad libitum. Experiments were performed during the day between 10:00 am and 2:00 pm. All experiments were performed according to the international norms of the ethical committee (DM42/2004-A).
.
Chemicals
Bovine serum albumin (BSA) and protein estimation kits were purchased from ZiestChem Company (Tehran, Iran). Kits for the estimation of urea, blood urea nitrogen (BUN), and creatinine were purchased from Sigma Company (St. Louis, MO, USA). Quercetin, 5,5-dithiobis(2-nitrobenzoic acid) [DTNB (Ellman’s reagent)], glacial acetic acid, heparin, nitroblue tetrazolium chloride (NBT), potassium dihydrogen phosphate (KH2PO4), reduced glutathione (GSH), sodium dihydrogen phosphate (NaH2PO4), NaF, trichloroacetic acid (TCA), thiobarbituric acid (TBA), and hydrogen peroxide were purchased from Sigma-Aldrich. All other chemical reagents and solvents were of analytical grade.
.
Animal Treatments
Animals were randomly divided into 5 groups of 10 animals each. Group I was kept as control group treated with 5% dimethyl sulfoxide for 7 consecutive days. Group II was treated with 5% dimethyl sulfoxide for 7 consecutive days followed by NaF (600 ppm) intoxication for the next 7 days. Groups III and IV were treated with quercetin (10 and 20 mg/kg) intraperitoneally for 7 days followed by NaF (600 ppm) in drinking water for the next 7 days. Group V was treated with the positive control, vitamin C (10 mg/kg body weight), intraperitoneally for 7 days followed by NaF (600 ppm) treatment for the same period.
.
Anesthesia and Tissue Collection
At the end of the treatment period, animals were anesthetized with ketamine (60 mg/kg) and xylazine (5 mg/kg) given intraperitoneally. Blood samples were collected from cardiac puncture and the kidneys were removed. Serums from the blood were separated by centrifugation at 12,000 × g for 10 min. All tissue samples were kept at −60°C until used for the biochemical studies.
.
Preparation of Tissue Homogenates
The whole kidney was homogenized (1:10 w/v) in phosphate-buffered saline (100 mM) containing ethylenediaminetetraacetic acid (1 mM, pH 7.4) and centrifuged (12,000 × g, 30 min, 4°C). The resulting supernatant was separated and used for the biochemical studies.
.
Determination of Protein Content
The protein content of rat kidney homogenates was determined by the method of Bradford using BSA as a standard.12
Biochemical Characterization
Estimation of lipid peroxidation
Lipid peroxidation as a measure of thiobarbituric acid reactive substances (TBARS) formation was determined by the method of Esterbauer and Cheeseman.13 Kidney tissue homogenates containing 1 mg protein was mixed with TCA (1 mL, 20%) and TBA (2 mL, 0.67%). The resulting reaction mixtures were incubated for 1 h at 100°C. After cooling, the precipitate was removed by centrifugation and the absorbance of the supernatant was measured at 535 nm using a blank containing all the reagents except the tissue homogenates.
.
Determination of superoxide dismutase activity
The reaction mixtures contained sodium carbonate (1 mL, 50 mM), NBT (0.4 mL, 25 μm), and freshly prepared hydroxylamine hydrochloride (0.2 mL, 0.1 mM). After mixing the reaction mixtures by inversion, the supernatants of kidney homogenates were added (containing 5 μg proteins). The change in absorbance of the reaction mixture was recorded at 560 nm.14
Determination of catalase activity
The catalase activity was measured by the method of Nabavi et al.1 Proteins contained in kidney tissue homogenates (5 μg) were mixed with hydrogen peroxide (2.1 mL, 7.5 mM), and a time scan was performed for 10 min at 240 nm at 25°C. The disappearance of peroxide, which reflects the level of catalase activity, was measured. One unit of catalase activity is defined as the amount of enzyme that reduces 1 μmol of hydrogen peroxide per minute.
Determination of reduced GSH activity
Reduced GSH level was determined according to the method of Ellman.15 Briefly, TCA (5%) was added to diluted kidney tissue homogenates (containing 36 μg protein) to precipitate the proteins. After centrifugation (12,000 × g, 5 min), the supernatant was taken and 5,5′-dithiobis(2-nitrobenzoic acid) solution (Ellman’s reagent) was added. The absorbance of the reaction mixture was measured at 417 nm. Various concentrations of reduced GSH were also used to construct a calibration curve from which the levels of reduced GSH in homogenate samples were calculated.
Serum biochemical parameters
BUN, creatinine, and serum urea concentration were assessed as markers of nephrotoxicity. All of these parameters were determined spectrophotometrically from serum samples by using commercially available kits and following the manufacturers’ procedures.
Statistical Analysis
All data are presented as means ± SD. Differences between group means were analyzed using one-way analysis of variance followed by Duncan’s multiple range tests. Results were considered statistically significant when p-value was <0.05.
.
RESULTS
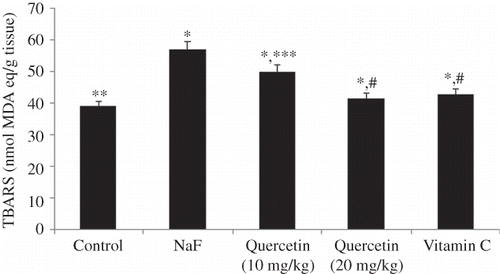
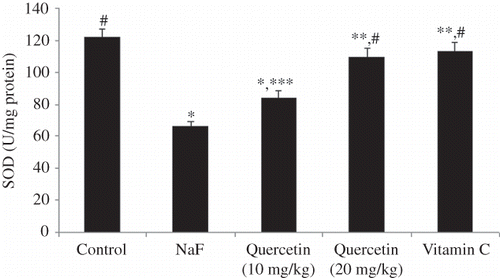
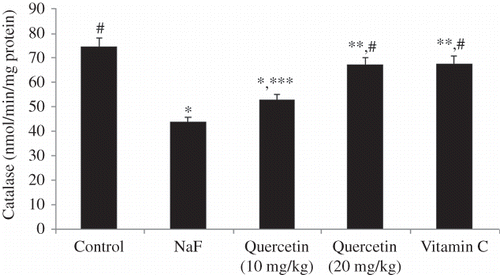
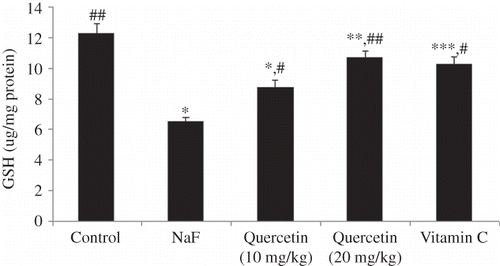
The level of lipid peroxidation in the kidney of all treatment groups is shown in Figure 1. A significant increase in lipid peroxidation of kidney tissues was observed when rats were intoxicated with NaF (46% vs. control, p < 0.001). Pretreatment of rats with quercetin for 7 days resulted in a dose-dependent reversal of the NaF-induced enhanced lipid peroxidation in the kidney. As shown in Figure 1, the positive control, vitamin C, administered at a dose of 10 mg/kg could also suppress the NaF-induced lipid peroxidation in the kidney. The effect of vitamin C was comparable with that of quercetin administered at a dose of 20 mg/kg (p > 0.05). The effect of quercetin on superoxide dismutase (SOD) activity of the kidney tissues is shown in Figure 2. Treatment of rats with NaF resulted in a 45.9% suppression of SOD activity (Figure 2). Quercetin treatment resulted in a dose-dependent reversal of the NaF-mediated amelioration of SOD activity. At the dose of 20 mg/kg, quercetin (p < 0.05) treatment as well as vitamin C (p < 0.05) could offer significant protection from NaF-mediated loss of SOD activity. As shown in Figure 3, treatment of rats with NaF resulted in a significant loss (41.5%, p < 0.001) of catalase activity. Although the NaF-mediated loss of catalase activity was not totally recovered by quercetin at the tested doses, a significant dose-dependent protection was observed (Figure 3). The activity profile of the positive control, vitamin C (p < 0.05), also mirrors that of quercetin administered at 20 mg/kg (p < 0.05). Figure 4 shows the reduced GSH levels in kidney tissue homogenates of different treatment groups. Exposure of rats to NaF caused a significant reduction (47.2%, p < 0.001) in reduced GSH level in the homogenates of rat’s kidney. As with the SOD and catalase activity studies, treatment with quercetin at 20 mg/kg and vitamin C resulted in a significant improvement of renal glutathione level following NaF exposure (p < 0.01). The serum biochemical parameters recorded for all the treatment groups are summarized in Table 1. The inclusion of NaF (600 ppm) in drinking water for 7 consecutive days resulted in a significant increase in serum creatinine (72.3%, p < 0.01), serum urea (39%, p < 0.001), and BUN (39%, p < 0.001) compared with normal (vehicle-treated) group. Pretreatment of rats with quercetin resulted in a dose-dependent improvement of all the biochemical parameters studied. As shown in Table 1, the activity profile of quercetin at 20 mg/kg was comparable with vitamin C at a dose of 10 mg/kg.
Figure 1. Effect of quercetin and vitamin C on TBARS level in sodium fluoride (NaF) intoxicated rat. Data are mean ± SD values (n = 10). Notes: *p < 0.001 versus control; **p > 0.05 versus control; ***p < 0.05 versus NaF; #p < 0.001 versus NaF.
Display full size
Figure 2. Effect of sodium fluoride (NaF) intoxication on superoxide dismutase (SOD) activity in the rat kidney. Data are mean ± SD values (n = 10). Notes: *p < 0.001 versus control; **p < 0.05 versus control; ***p < 0.01 versus NaF; #p < 0.001 versus NaF.
 Display full size
Display full size
Figure 3. Effect treatment on catalase levels in sodium fluoride (NaF) intoxicated rat. Data are mean ± SD values (n = 10). Notes: *p < 0.001 versus control; **p < 0.05 versus control; ***p < 0.05 versus NaF; #p < 0.001 versus NaF.
 Display full size
Display full size
Figure 4. Toxic effect of sodium fluoride (NaF) on levels of reduced glutathione (GSH) in the kidney homogenate of rat. Data are mean ± SD values (n = 10). Notes: *p < 0.001 versus control; **p < 0.05 versus control; ***p < 0.01 versus control; #p < 0.01 versus NaF; ##p < 0.001 versus NaF.
 Display full size
Display full size
Table 1. Effect of quercetin on serum creatinine, serum urea, and blood urea nitrogen (BUN) levels in sodium fluoride-induced nephrotoxicity in rats.
CSV Display Table
DISCUSSION
Several studies have demonstrated that the toxicity of fluoride intoxication is associated with excessive generation of ROS, a reduction in antioxidant defenses including reduced catalase, and SOD activity coupled with a diminished level of reduced GSH.16 It has also been reported that fluoride intoxication results in excessive generation of reactive nitrogenous species such as nitric oxide (NO).16 All these altered oxidant–antioxidant balance will inevitably lead to lipid peroxidation and damage to macromolecular molecules such as cell membrane, DNA, and proteins.16 At the molecular level, fluoride ions have also been shown to directly inhibit glycolysis by inhibiting the enzyme enolase. In renal cells, glycolysis inhibition is often compensated by using alternative energy-producing pathways that lead to cellular dysfunctions.17 Some studies further suggest that NO is implicated to the fluoride-mediated renal dysfunction,17 and therapeutic intervention by iNOS inhibitors has been proposed.18 On the basis of these findings, this study was designed to instigate the extent of fluoride-mediated alteration of oxidant–antioxidant balance in the kidney and whether a beneficial in vivo effect is demonstrated to one of the most popular antioxidant polyphenol, quercetin.
In order to investigate the vast arrays of changes to oxidant–antioxidant status of the kidney following fluoride administration, our experimental protocol was designed to include measurements of TBARS, the activity of the antioxidant enzymes (SOD and catalase), and the level of the cellular GSH and serum biochemical markers including BUN, urea, and creatinine. In agreement with previous studies, our study clearly demonstrated that fluoride causes significant rat kidney damage when administered for 7 days via drinking water (600 ppm). As assessed by the measurement of SOD and catalase activities as well as GSH and the lipid peroxidation levels, antioxidant defenses significantly diminished following fluoride intoxication. Quercetin treatment at doses of 10 and 20 mg/kg body weight prior to fluoride intoxication restored the level/activity of antioxidant defenses in rat kidney in a dose-dependent manner. When an antioxidant positive control, vitamin C at a dose of 10 mg/kg, was used before fluoride intoxication, similar restoration of antioxidant defenses was achieved. Our result for the first time therefore provides direct evidence on the potential of antioxidant compounds in specifically reversing prooxidative kidney damage induced by fluoride. In addition to the direct effect of fluoride on oxidant–antioxidant balance in the kidney, the serum levels of BUN, creatinine, and urea were measured following fluoride administration. As with previous reports,19 our results also revealed augmented serum level of markers, suggesting glomerular damage by fluoride intoxication. The administration of quercetin and vitamin C once again reversed the serum markers of glomerular damage, further substantiating the beneficial effect of antioxidant compounds.
In addition to NO,16 ROS, including hydroxyl radicals, have been implicated in the initiation and/or progression of nephrotoxicity.20 On the other hand, transition metals such as iron ion generate hydroxyl radicals and other ROS through the Fenton reaction.19 As an efficient reactive oxygen scavenger including NO and iron chelator, it appears that quercetin improves the oxidant–antioxidant imbalance created by fluoride intoxication.21 As a multifunctional molecule, however, other mechanisms of action are also likely to be associated with quercetins nephroprotective effect.
CONCLUSION
This study provided direct evidence for the altered antioxidant–oxidant balance in the kidney following fluoride intoxication. This study also demonstrated the direct beneficial effect of quercetin in preventing renal damage induced by fluoride. On the basis of the data provided in this study, further studies on clinical trial and toxicological parameters of quercetin and related natural products are well merited.
ACKNOWLEDGMENTS
The authors acknowledge the financial support of National Elite’s Foundation of Iran (Tehran, Iran) for this study. This article is dedicated to Seyed Maryam Nabavi and the memory of Seyed Ali Asghar Nabavi.
REFERENCES
-
Nabavi SF, Moghaddam AH, Eslami S, Nabavi SM. Protective effects of curcumin against sodium fluoride-induced toxicity in rat kidneys. Biol Trace Elem Res. 2012;145:369–374.[Crossref], [PubMed], [Web of Science ®], [Google Scholar]
-
World Health Organization. Environmental Health Criteria, 227 Fluorides IPS, Geneva, Switzerland; 2002.[Google Scholar]
-
Patel PD, Chinoy NJ. Influence of fluoride on biological free radical reactions in ovary of mice and its reversal. Fluoride. 1998;31:S27.[Web of Science ®], [Google Scholar]
-
Xiong X, Liu J, He W, . Dose effect relationship between drinking water fluoride levels and damage to liver and kidney functions in children. Environ Res. 2007;103:112–116.[Crossref], [PubMed], [Web of Science ®], [Google Scholar]
-
Kobayashi CAN, Leite AL, Silva TL, . Proteomic analysis of kidney in rats chronically exposed to fluoride. Chem-Biol Interact. 2009;180:305–311.[Google Scholar]
-
Urquiaga I, Leighton F. Plant polyphenol antioxidants and oxidative stress. Biol Res. 2000;33:55–64.[Crossref], [PubMed], [Web of Science ®], [Google Scholar]
-
Harborne JB. Plant phenolics. In: Bell EA, Charlwood BV, eds. Encyclopedia of Plant Physiology, Volume 8, Secondary Plant Products. Berlin, Heidelberg, New York: Springer-Verlag; 1980:329–395.[Google Scholar]
-
Dias AS, Porawski M, Alonso M, Marroni N, Collado PS, Gonzalez-Gallego J. Quercetin decreases oxidative stress, NF-kB activation, and iNOS overexpression in liver of streptozotocin-induced diabetic rats. J Nutr. 2005;135:2299–2304.[Google Scholar]
-
Behling EB, Sendão MC, Francescato HDC, Antunes LMG, Costa RS, Bianchi MLP. Comparative study of multiple dosage of quercetin against cisplatin-induced nephrotoxicity and oxidative stress in rat kidneys. Pharmacol Rep. 2006;58:526–532.[PubMed], [Web of Science ®], [Google Scholar]
-
Sánchez-González P, López-Hernández FJ, Prierto M, Vicente-Vicente L, López-Novoa JM, Morales AI. Renoprotective effects of quercetin on cisplatin-induced nephrotoxicity: Role of TNF-alpha, NF-kappaB and iNOS. Toxicol Lett. 2010;196:S242.[Google Scholar]
-
Francescato HD, Coimbra TM, Costa RS, Bianchi Mde L. Protective effect of quercetin on the evolution of cisplatin-induced acute tubular necrosis. Kidney Blood Press Res. 2004;27:148–158.[Crossref], [PubMed], [Web of Science ®], [Google Scholar]
-
Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254.[Crossref], [PubMed], [Web of Science ®], [Google Scholar]
-
Esterbauer H, Cheeseman KH. Determination of aldehydic lipid peroxidation products: Malonaldehyde and 4-hydroxynonenal. Method Enzymol. 1990;186:407–421.[Crossref], [PubMed], [Web of Science ®], [Google Scholar]
-
Nabavi SM, Nabavi SF, Eslami S, Moghaddam AH. In vivo protective effects of quercetin against sodium fluoride-induced oxidative stress in the hepatic tissue. Food Chem. 2012; 132:931–935.[Crossref], [Web of Science ®], [Google Scholar]
-
Ellman GL. Tissue sulphydryl group. Arch Biochem Biophys. 1959;82:70–77.[Crossref], [PubMed], [Web of Science ®], [Google Scholar]
-
Yu RA, Xia T, Wang AG, Chen XM. Effects of selenium and zinc on renal oxidative stress and apoptosis induced by fluoride in rats. Biomed Environ Sci. 2006;19:439–444.[Google Scholar]
-
Goligorsky MS, Brodsky SV, Noiri E. Nitric oxide in acute renal failure: NOS versus NOS. Kidney Int. 2002;61:855–861.[Crossref], [PubMed], [Web of Science ®], [Google Scholar]
-
Guven A, Uysal B, Akgul O, . Scavenging of peroxynitrite reduces renal ischemia/reperfusion injury. Ren Fail. 2008;30:747–754.[Taylor & Francis Online], [Web of Science ®], [Google Scholar]
-
Nabavi SF, Nabavi SM, Moghaddam AH, Naqinezhad A, Bigdellou R, Protective MS. Protective effects of Allium paradoxum against gentamicin-induced nephrotoxicity in mice. Food Funct. 2012;3:28–29.[Crossref], [PubMed], [Web of Science ®], [Google Scholar]
-
Singh P, Srivastava MM, Khemani LD. Renoprotective effects of Andrographis paniculata (Burm. f.) Nees in rats. Upsala J Med Sci. 2009;114:136–139.[Taylor & Francis Online], [Google Scholar]
Nabavi SF, Nabavi SM, Abolhasani F, Moghaddam AH, Eslami S. Cytoprotective effects of curcumin on sodium fluoride-induced intoxication in rat erythrocytes. Bull Environ Cont Toxicol. 2012;88:486–490.[Crossref], [PubMed], [Web of Science ®], [Google Scholar]
